卵成熟過程における細胞骨格間クロストーク
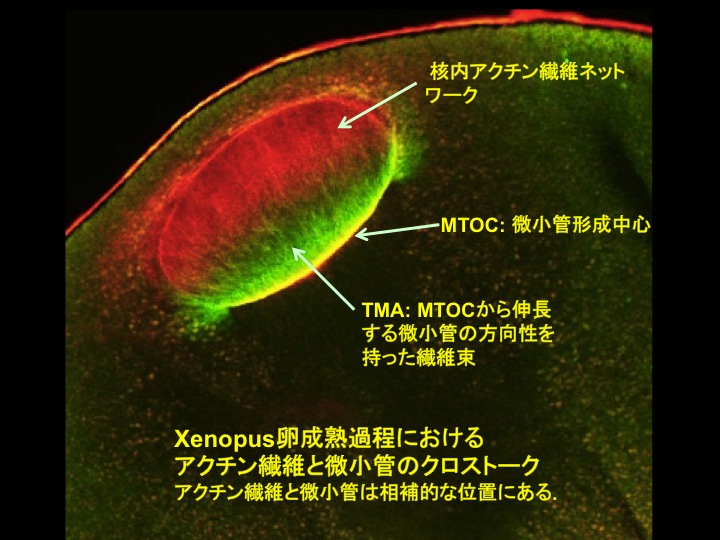
近年, 紡錘体の形成や卵表層への移動にアクチン繊維が関与することが種々の動物卵で見出され, 紡錘体とアクチン繊維との関係に注目が集まっている. また, 我々は独自に, 核膜を裏打ちしている中間径繊維のカテゴリーに含まれるラミン繊維の分散にはアクチン繊維の再編成が重要であることを見出し報告した. このように細胞骨格の相互作用は複雑に絡み合い, その総体として核膜の分散や紡錘体形成が進行すると考えられる. つまり, 問うべき主題は, 卵成熟の進行に伴い, 3つの異なる細胞骨格である微小管・アクチン繊維・中間径繊維はどのような分子機構で相互作用して紡錘体形成に至るのか, ということである.
アクチン調節蛋白質コフィリンを核とする細胞骨格制御
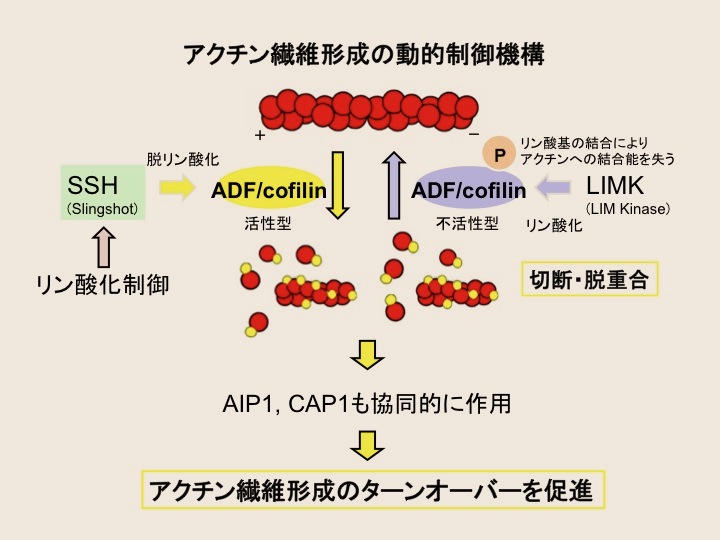
アクチン繊維を切断・脱重合するアクチン調節蛋白質コフィリンのリン酸化制御機構の存在が、卵成熟過程のアクチン繊維形成の動的な制御に必須であることを我々は報告してきた。コフィリンの作用を中心とするアクチン繊維の再編成は細胞機能にとって必須であり、そのリン酸化制御機構の詳細や、コフィリンと共同してアクチン繊維形成のダイナミクスを高める因子であるAip1やCAP1などの機能について、液-液相分離の観点も含め多角的に解析を進めている。
核内アクチン動態とヘテロクロマチン構築の関連
近年、核内アクチン繊維形成の動態がゲノムの転写や複製、遺伝子発現の初期化に関与しているという報告がなされ、その機能に注目が集まっている。そして2018年には、DNAの2本鎖切断の修復にArp2/3 complexを主体とするde novoのアクチン繊維形成が必須であるという予期せぬ結果が示された。我々は独自に、核内アクチン繊維形成の動態制御がヘテロクロマチン構造の変化と相関する現象を見出し、現在その分子機構の解析を行っている。

