神経細胞の成長円錐におけるlasp-2の挙動
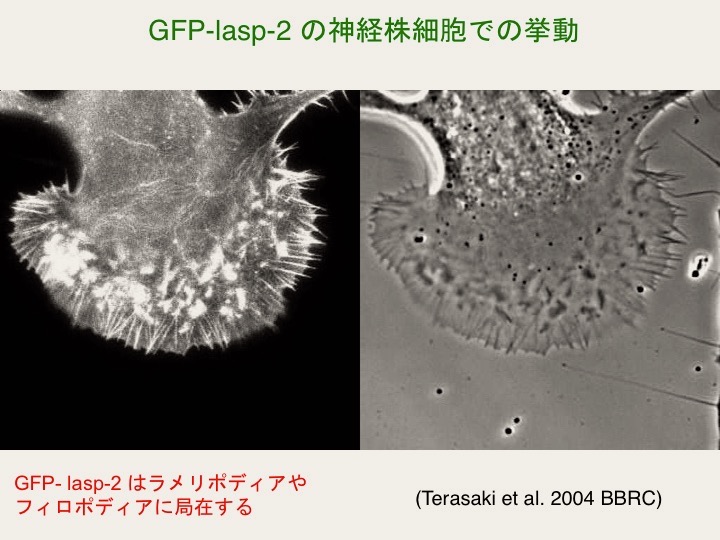
神経芽腫細胞株 NG108 において lasp-2 は成長円錐のフィロポディアやラメリポディアに局在する事を確認しています(Terasaki et al., 2004)。現在はニワトリの感覚神経細胞(脊髄後根神経節 = DRG)由来の初代培養細胞を用いて成長円錐の接着構造やエンドソームにおける lasp-2 の挙動を観察するとともにラットの海馬の神経細胞のスパインにおける lasp-2 の局在も解析しています。

本研究室ではニワトリの脳から発見した新しいアクチン結合タンパク質 lasp-2(LIM and SH3 protein 2)の神経細胞やグリア細胞における機能や活性制御機構、分子進化を調べています。神経細胞のネットワーク形成では軸索(axon)の先端部の成長円錐(growth cone)が運動を続けて標的に到達し、シナプスの送り手側は神経終末、受け手側はスパイン(spine)となります。成長円錐やスパインの形成にはアクチン結合タンパク質が必須ですが、神経細胞におけるアクチン結合タンパク質の含量は筋肉などに比べて少ないため生化学的な解析は難しいとされてきました。しかし当研究室ではFアクチンカラムでニワトリ脳抽出液を解析して lasp-2 を見つける事ができました。Lasp-2 が哺乳類や鳥類の脳で高発現している事から神経細胞とグリア細胞(アストロサイト)を主に用いて機能解析を進めています。
神経芽腫細胞株 NG108 において lasp-2 は成長円錐のフィロポディアやラメリポディアに局在する事を確認しています(Terasaki et al., 2004)。現在はニワトリの感覚神経細胞(脊髄後根神経節 = DRG)由来の初代培養細胞を用いて成長円錐の接着構造やエンドソームにおける lasp-2 の挙動を観察するとともにラットの海馬の神経細胞のスパインにおける lasp-2 の局在も解析しています。
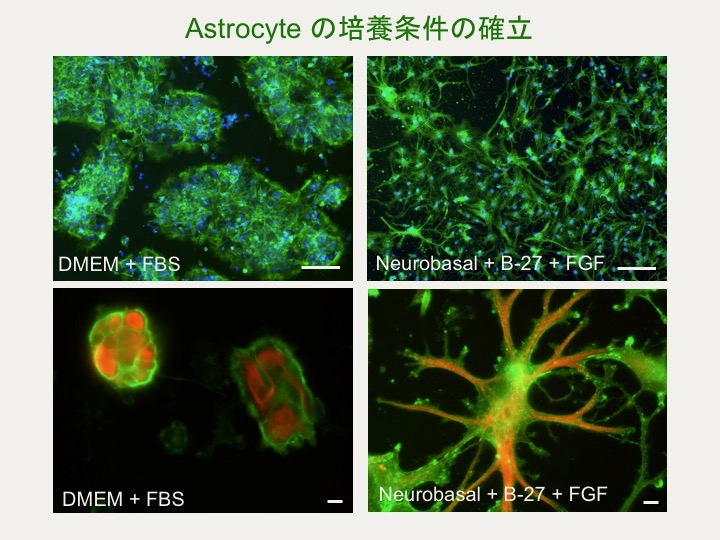
中枢神経のグリア細胞であるアストロサイトは放射状に伸びた終足の末端のフィロポディアを介して神経細胞に連結し、シナプスの情報伝達などを制御しますが、アクチン系細胞骨格の研究は十分に行われていません。 そこでニワトリ大脳由来のアストロサイトを用いて lasp-2 の挙動を解析しています。その過程でニワトリアストロサイトが生体内に近い突起を形成する培養条件や凍結技術も確立しました。
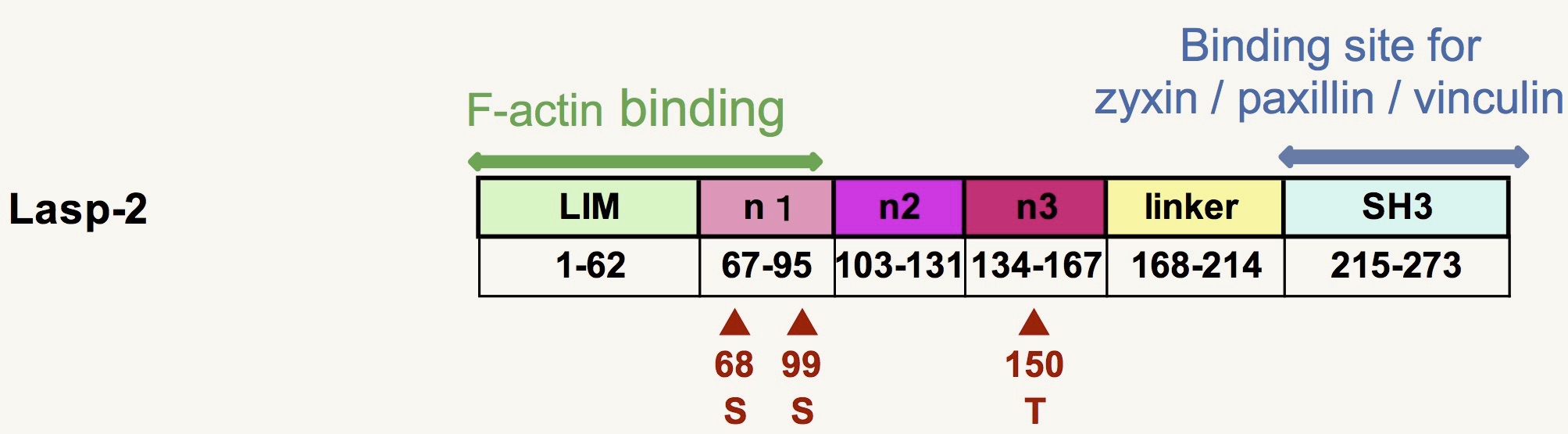
Lasp-2 の細胞内局在とアクチン結合活性には LIM ドメインと最初の nebulin リピートが必須である事や(Nakagawa et al., 2009)、lasp-2がリン酸化を受ける事を確認しました。現在はリン酸化が予測される部位の変異体を作成して 局在やアクチン結合活性を解析中です。また、lasp-2 アフィニティカラムを用いて lasp-2 結合タンパク質の探索も進めています。
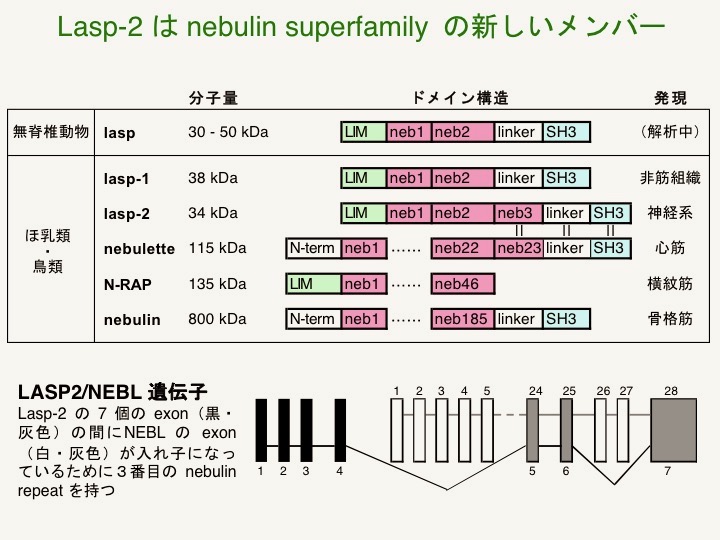
ほ乳類と鳥類でnebulin リピートを持つタンパク質は、lasp-1・lasp-2・nebulette・N-RAP・nebulinの5種で、nebulette は lasp-2 の遺伝子に exon が挿入されたアイソフォームである事や(Terasaki et al., 2006)、ホヤ laspはlasp-1 と lasp-2 の機能を兼ねていることを明らかにしました(Terasaki et al., 2008)。現在は様々な生物のnebulin superfamilyのシンテニー(ゲノム上の位置関係の相同性)などから分子進化の過程を推測しています。
研究指導の目標は論理的思考・技術に対する正しい理解・データに対する責任を身につけることです。
〒263-8522
千葉県千葉市稲毛区弥生町 1-33
●JR西千葉駅より徒歩2分
●京成みどり台駅より徒歩7分
●モノレール天台駅より徒歩10分
