同形胞子シダ植物における隠蔽種探索
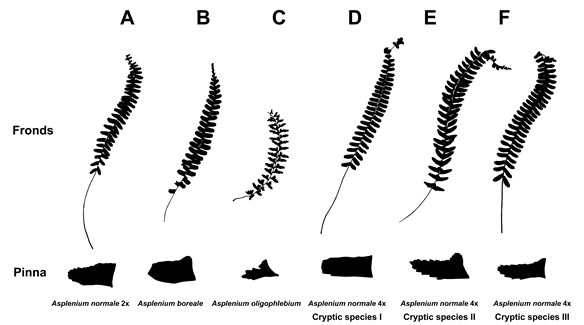
生殖的隔離を持つ別種でありながら、形態的識別が困難であるがゆえに従来同じ種にまとめられていたものを隠蔽種と呼びます。隠蔽種は種分化して間もない発端種である可能性が高く、種分化機構の解明に適した材料です。シダ植物は花を持たず形態的に単純であるため、潜在的な隠された種多様性が存在する可能性があります。今迄のところ、ミズワラビ(1, 3)、フモトシダ、ヌリトラノオ(9)、ノキシノブ(19)に隠蔽種が含まれていることを明らかにしました。これらは全て4倍体であり、倍数化が隠蔽種を産みだすパターンが見えてきました。
同形胞子シダ植物における交雑種分化

交雑種分化とは種間交雑が新しい種を生み出すタイプの種分化です。シダ植物で見られるのは、高次倍数化を伴う交雑種分化です。一つは無融合生殖種と有性生殖種との交雑により、新たな無融合生殖クローンが生み出されるパターン。これは、無融合生殖種が精子を作る能力を保持しており、無融合生殖の特性が優性(顕性)遺伝することが原因となっています。もう一つは有性種間の不稔雑種が倍数化によって稔性を回復する異質倍数体形成と呼ばれる種分化です。無融合生殖種であるオオバノイノモトソウは、前者の機構で多様なクローンを形成しています(7)。後者の異質倍数体形成はテーマ1の隠蔽種と強く関連しています。
異質倍数体の独立複数回起源は種分化につながるか?
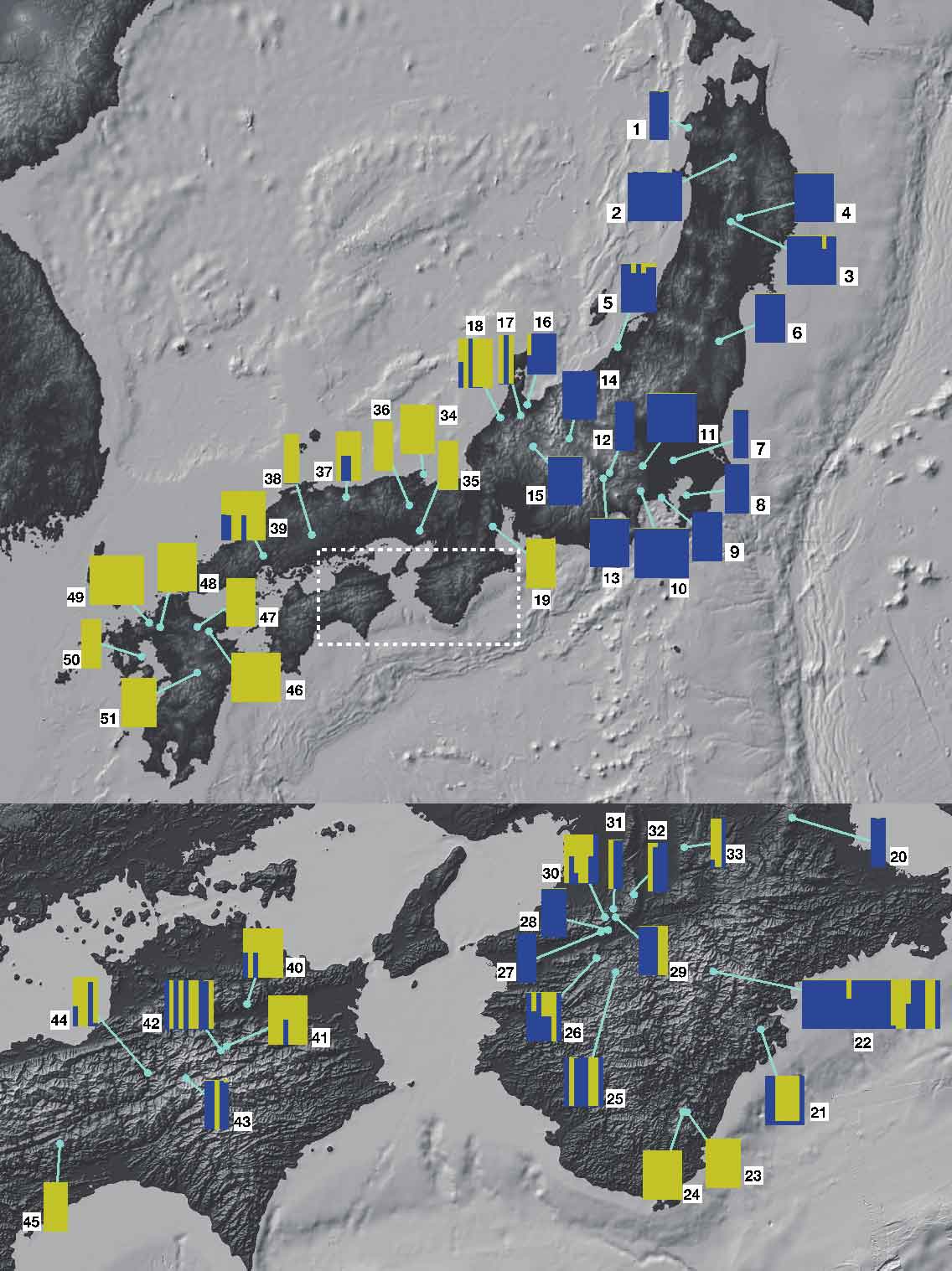
異質倍数体形成は同じ祖先種の組み合わせから、何度も繰り返し生じることが多い事が知られています。独立に生じた系統は融合して一つの種となる場合もあれば、生殖的隔離を発達させて別種となる可能性も指摘されています。ノキシノブから分離されて新種記載されたクロノキシノブでは、日本列島内での東西での遺伝的分化が見られます。東西のタイプ間の雑種は部分的に不稔です。おそらく、地理的に異なる地域で独立に生じたものが、分布を拡大し、今まさに近畿を中心に出会ったところではないかと考えています。内在的生殖隔離が独立起源の異質倍数体系統で急速に進化する可能性を示しています。
ハイマツ・キタゴヨウ間の浸透性交雑現象の解析

浸透性交雑とは,遺伝的に分化した分類群が二次的に接触して交雑し,遺伝的交流を行う現象を言います。生物学的種概念によれば,“種とは遺伝的に隔離されているもの”と定義されます。従って浸透性交雑を行う種群は,“種とは何か”という古くて新しい問題にアプローチする際に,最適な実験系であると考えています。母性遺伝のミトコンドリアと父性遺伝の葉緑体という変わったオルガネラ遺伝様式を持つマツ科植物に着目し、ハイマツとキタゴヨウという標高によってすみ分ける二種の間の交雑帯の遺伝的構造を明らかにしています(2, 4, 5)。

